

Neueste Studien über Schlankkräuselspinnen
Die in den Tropen Südostasiens mit 57 Arten verbreiteten Schlankkräuselspinnen (Psechridae) wurden vom derzeitigen Volontär in der Zoologie Steffen Bayer im Rahmen seiner Doktorarbeit bearbeitet.
Bis Ende des letzten Jahrzehnts war über die erst 1890 aufgestellte Familie der Psechridae nur wenig bekannt. Die ersten Arten waren erstmals 1857 ursprünglich als Winkelspinnen beschrieben worden. Die beiden Gattungen der Familie Psechrus (Kräuseldeckennetzspinnen) und Fecenia (Pseudo-Radnetzspinnen) wurden erst später aufgestellt.
Merkmale, anhand derer man die einzelnen Arten unterscheiden kann (diagnostische Merkmale) waren vielfach noch nicht verstanden und auch sonst war z.B. über die Lebensweise der meisten Arten nur wenig bekannt.
Einige der Typus-Exemplare der einzelnen Arten sind noch nicht ausgewachsene Tiere, bei denen die artspezifischen Begattungsorgane noch nicht voll ausgebildet waren. Dies erschwerte zusätzlich die Charakterisierung der einzelnen Arten. Typus-Exemplare sind die individuellen Tiere (in diesem Falle also tote, konservierte Spinnen), anhand derer eine bestimmte Art erstmals beschrieben wurde. Der Typus stellt also den Namensbezug zu der entsprechenden Art her und stellt somit den „Urmeter“ für alle Artgenossen dar.
Oftmals wurde von früheren Autoren auch die Artenvielfalt der Psechriden unterschätzt. Gründe dafür waren fehlende oder unzureichende gezielte Aufsammlungen dieser Tiere sowie fehlende Recherchen in den Sammlungen der naturwissenschaftlichen Museen weltweit. Nicht selten wurden auch klare Merkmalsunterschiede, anhand welcher man verschiedene Arten hätte abgrenzen können (interspezifische Merkmalsunterschiede), als weniger bedeutend angesehen und als intraspezifische Variation betrachtet. All dies erschwerte die exakte Bestimmung von Schlankkräuselspinnen.
Seit 2009 beschäftigt sich Steffen Bayer mit der Taxonomie (Artenforschung) sowie der molekularen Phylogenie (Arten-Verwandtschaftsforschung) der Schlankkräuselspinnen. Aufgrund der Ergebnisse seiner Studien, die z.T. in Kooperation mit anderen Wissenschaftlern erarbeitet wurden, konnten die Psechriden eingehend charakterisiert werden. Die Vertreter der artenreichen Gattung Psechrus wurden in Artengruppen aufgeteilt, insgesamt 30 neue Arten erstmals beschrieben, jeweils ein Bestimmungsschlüssel für beide Gattungen vorgelegt und die Verwandtschaftsverhältnisse innerhalb dieser Spinnengruppe sowie zu anderen Spinnengruppen aufgezeigt (Bayer 2011, 2012, 2014; Bayer & Jäger 2010, Bayer & Schönhofer 2013).
Taxonomie und systematische Einordnung
Die Stellung der Psechridae im System der Webspinnen
Aufgrund des Besitzes von Cribellum und Calamistrum wurden die Psechriden zu Beginn des 20. Jahrhunderts zur damals als monophyletisch erachteten Großgruppe der Cribellatae gerechnet und dort meist als nächste Verwandte der Uloboriden angesehen. Morphologisch-kladistische Untersuchungen, worin Psechriden einbezogen wurden, gab es ab Anfang der 1990er Jahre (Griswold 1993, Griswold et al. 2005, Silva 2003). Die Psechriden wurden hierbei immer der Überfamilie Lycosoidea zugeordnet. Eine nahe Verwandtschaft mit den Senoculiden wurde als wahrscheinlich betrachtet. In einer molekulargenetisch-phylogenetischen Studie, bei welcher die beiden Markergene 28S rRNA und COI analysiert wurden (Bayer & Schönhofer 2013), konnte neuerdings die Zugehörigkeit von Psechrus und Fecenia zu den Lycosoidea bestätigt werden.
Die Beziehungen zu den anderen Familien dieser Überfamilie blieben allerdings auch nach dieser Studie unklar. Jedoch konnten die Psechrus-Artengruppen, die anhand morphologischer Merkmale aufgestellt wurden, bestätigt werden, d.h. als Monophyla erkannt werden.
Lebensraum und Lebensweise
Die nachtaktiven Schlankkräuselspinnen kommen in tropischen, immergrünen Wäldern Südostasiens vor. Offene Lebensraumtypen werden zumeist gemieden. Kräuseldeckennetzspinnen (Psechrus) sind meist in Bodennähe zu finden und spannen ihre horizontalen, leicht gewölbten Gewebedecken, an deren Unterseite sie sich blitzschnell bewegen, oft zwischen Baumwurzeln, Felsen, umherliegenden Steinbrocken oder unterhalb sandiger Überhänge auf.
Pseudoradnetzspinnen (Fecenia) hingegen sind typische Bewohner der Laubschicht und legen vertikale Pseudoradnetze an, in die stets ein eingerolltes Blatt integriert ist, welches der Spinne als Schutzbehausung dient. Diese Netze finden sich selbst noch im Baumkronenbereich. Daher besteht die Beute von Fecenia vorrangig aus nachtaktiven Fluginsekten (z.B. Nachtfaltern). Kräuseldeckennetzspinnen erbeuten neben Fluginsekten auch laufende Insekten, wie z.B. Schaben oder Grillen, die auf die Netzdecke fallen.
Beide Gattungen gehören also zu den cribellaten Spinnen und haben durch ihre äußerst effektive Fangwolle die Möglichkeit, auch Beutetiere zu überwältigen, die größer sind als sie selbst. Die langwierige Herstellung dieser Fäden sowie Ausbesserungen erfolgen stets in der zweiten Nachthälfte.
Paarungsbereite Männchen finden die Netze von Weibchen höchstwahrscheinlich anhand von Pheromonen (Sexuallockstoffe), die von diesen in die Fäden des Netzes integriert wurden. Ist ein weibliches Netz erreicht, beginnt ein recht langes Repertoire an verschiedenen Zupfbewegungen, die dem Weibchen signalisieren, dass sich ein Geschlechtspartner nähert und nicht etwa Beute. Der eigentlichen Kopulation geht stets ein Vorspiel voraus, in welchem die Beine des Weibchens zart gestreichelt werden. Bei den Pseudoradnetzspinnen kommt es vorher noch zu einer recht langen Phase, in der das Männchen auf dem eingerollten Blatt des Weibchens sitzt und dieses auf spezielle Art beklopft. Die Kopulation findet dann nachts in der Mündung des eingerollten Blattes statt. Danach sitzen beide Partner noch einige Zeit zusammen im eingerollten Blatt. Im Falle von Psechrus findet die Kopulation unterhalb der Gewebedecke statt, in aller Regel ebenfalls nachts. Sie dauert mitunter nur wenige Sekunden.


Färbung und Zeichnungsmuster
Die Färbung der Schlankkräuselspinnen variiert nur geringfügig und ist generell (hell) bräunlich oder gelblich. Der grundlegende Unterschied im Zeichnungsmuster der beiden Gattungen besteht darin, dass bei den Kräuseldeckennetzspinnen (Psechrus) ventral am Opisthosoma (Hinterleib) eine helle, relativ dünne Medianlinie und bei den Pseudoradnetzspinnen (Fecenia) zwei benachbarte, parallel zueinander orientierte, längliche, helle Flecken erkennbar sind (Bayer 2011). Auch andere Aspekte des Zeichnungsmusters variieren innerhalb der beiden Gattungen nur wenig. Bei den Kräuseldeckennetzspinnen gibt es Artengruppen-spezifische Unterschiede in der Breite der oben angesprochenen Medianlinie sowie der lateralen, dunklen Bänder dorsal auf dem Prosoma (Vorderleib) (Bayer 2012). In wenigen Fällen kann auch die Zeichnung auf dem Sternum (Brustschild) Artengruppen-spezifisch sein. Was die Pseudoradnetzspinnen anbelangt, gibt es praktisch keine Zeichnungsaspekte, die artspezifisch sind.
Taxonomische Bedeutung von Prä-Epigyne und Prä-Vulva
Subadulte Weibchen, also solche, welche lediglich eine Häutung durchlaufen müssen, um geschlechtsreif zu werden, besitzen bereits eine Vorläuferstruktur an der Stelle, an welcher sich bei geschlechtsreifen (adulten) Weibchen das Kopulationsorgan befindet (außenliegender Teil wird als »Epigyne« bezeichnet, von außen nicht sichtbarer Teil »Vulva«). Dementsprechend wird die Struktur bei einem subadulten Weibchen als Prä-Epigyne bzw. Prä-Vulva bezeichnet. Diesen wurde bislang in arachnologischen Studien kaum Beachtung geschenkt. Mithilfe spezieller Präparationsmethoden konnte nun für die Psechriden interessanterweise herausgefunden werden, dass Prä-Epigyne und -Vulva sogar artspezifisch sind. Für die Gattung Fecenia konnten die Prä-Epigynen-Merkmale sogar erstmals in der Geschichte der Arachnologie, in einen Arten-Bestimmungsschlüssel integriert werden (Bayer 2011).
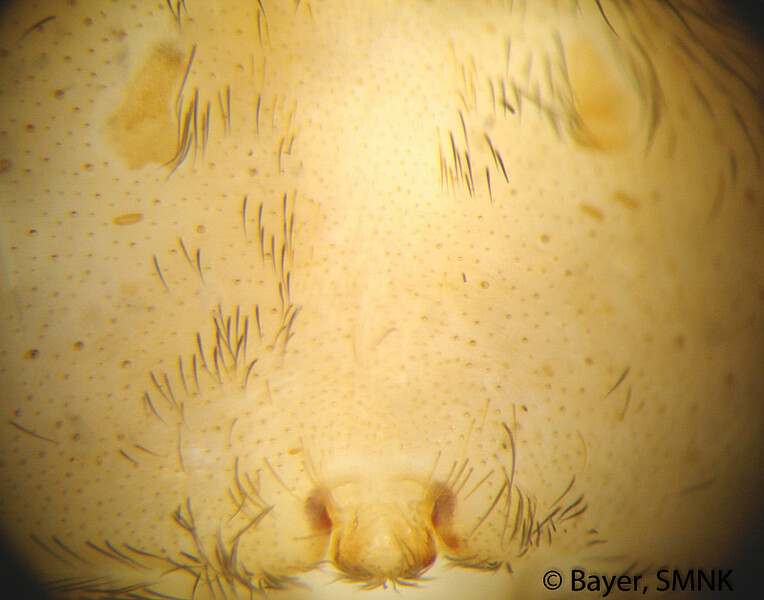
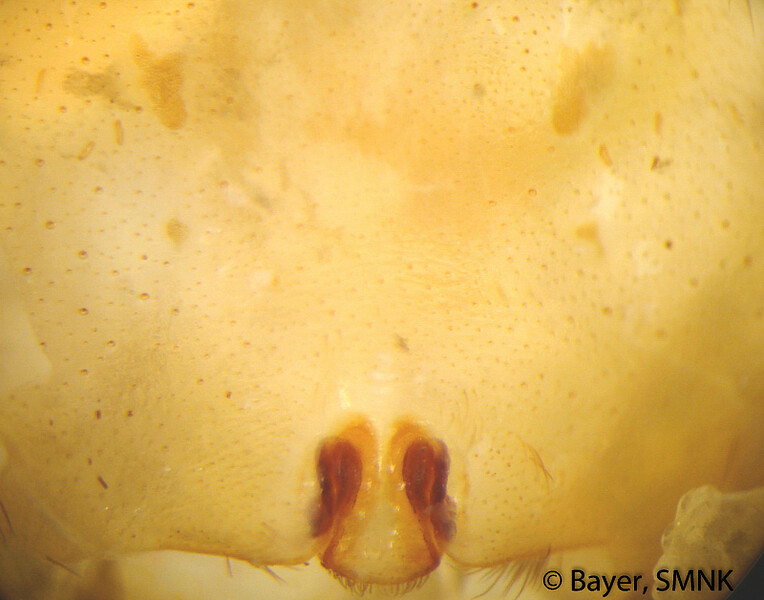
Arten
Gattung Psechrus
argentatus-group
P. argentatus (Doleschall, 1857)
P. libelti Kulczyński, 1908
mulu-group
P. mulu Levi, 1982
P. borneo Levi, 1982
P. ulcus Bayer, 2012
P. kinabalu Levi, 1982
annulatus-group
P. annulatus Kulczyński, 1908
P. aluco Bayer, 2012
P. decollatus Bayer, 2012
singaporensis-group
P. singaporensis Thorell, 1894
P. elachys Bayer, 2012
P. norops Bayer, 2012
P. arcuatus Bayer, 2012
ancoralis-group
P. ancoralis Bayer & Jäger, 2010
P. rani Wang & Yin, 2001
P. laos Bayer, 2012
P. antraeus Bayer & Jäger, 2010
P. khammouan Jäger, 2007
P. steineri Bayer & Jäger, 2010
himalayanus-group
P. himalayanus Simon, 1906
P. marsyandi Levi, 1982
P. inflatus Bayer, 2012
P. ghecuanus Thorell, 1897
P. pakawini Bayer, 2012
P. luangprabang Jäger, 2007
P. demiror Bayer, 2012
P. arietinus Bayer, 2014
P. jaegeri Bayer, 2012
P. vivax Bayer, 2012
P. insulanus Bayer, 2014
sinensis-group
P. sinensis Berland & Berland, 1914
P. triangulus Yang, Zhang, Zhu & Song, 2001
P. tingpingensis Yin, Wang & Zhang, 1985
P. obtectus Bayer, 2012
P. fuscai Bayer, 2012
P. kunmingensis Yin, Wang & Zhang, 1985
P. jinggangensis Wang & Yin, 2001
P. ampullaceus Bayer, 2014
P. senoculatus Yin, Wang & Zhang, 1985
P. kenting Yoshida, 2009
P. taiwanensis Wang & Yin, 2001
P. clavis Bayer, 2012
torvus-group
P. torvus (O. Pickard-Cambridge, 1869)
P. hartmanni Bayer, 2012
P. zygon Bayer, 2012
P. tauricornis Bayer, 2012
Incertae sedis (Arten, die keiner, der bislang aufgestellten Artengruppen zugeordnet werden konnten)
P. crepido Bayer, 2012
P. omistes Bayer, 2014
P. quasillus Bayer, 2014
P. huberi Bayer, 2014
P. schwendingeri Bayer, 2012
P. wade Bayer, 2014
P. cebu Murphy, 1986
Gattung Fecenia
F. ochracea (Doleschall, 1859)
F. macilenta (Simon, 1885)
F. protensa Thorell, 1891
F. cylindrata Thorell, 1895


")
, vereinfacht (Grafik: S. Bayer)")